Controlling Endotoxin Contamination in Bioprocessing


Scientists working with cell cultures or relying on recombinant proteins and other biological products are familiar with the perils of endotoxin contamination. Endotoxins are bacterial cell-associated toxins and represent a significant contaminant of concern in maintaining optimal cell culture conditions and producing high-quality and safe biologicals. For example, during bioprocessing, endotoxins originating from bacterial contamination may impact the properties of cells in bioreactors, thereby detrimentally affecting production yields (Epstein et al. 1990). Additionally, biological therapeutics and research reagents contaminated with endotoxins lead to adverse reactions in patients and induce unexpected toxicities and unwanted activities in preclinical settings. Therefore, a great deal of effort is devoted to preventing and removing endotoxin contamination in bioprocessing workflows.
What are endotoxins?
Endotoxin refers to a bacterial cellular component with associated toxicity. Specifically, it refers to lipopolysaccharide (LPS), a significant constituent of the Gram-negative bacterial (e.g., Escherichia coli, Klebsiella, and Pseudomonas) cell wall’s outer membrane. Unlike exotoxins, which are toxic proteins secreted by live bacteria, endotoxin release occurs upon bacterial death or lysis.
Structurally, LPS consists of two main organic molecules; a hydrophobic Lipid A that anchors LPS to the outer membrane and a complex polysaccharide tail. The Lipid A moiety is a bisphosphorylated diglucosamine backbone covalently linked to multiple acyl chains of variable lengths (Ohto et al. 2012, Su and Ding, 2015). The carbohydrate component is attached to one Lipid A glucosamine group and consists of a core polysaccharide and O-specific repeating oligosaccharides or O-antigen.

Reprinted from "Gram negative bacteria cell wall structure" by BioRender.com (2021). Retrieved from https://app.biorender.com/biorender-templates.
The Gram-negative bacterial envelope consists of an inner membrane, periplasmic peptidoglycan layer, and outer membrane. The endotoxin, Lipopolysaccharide (LPS), is a major component of the outer leaflet of the Gram-negative bacterial outer membrane (Sampath 2018).
Basis of LPS toxicity
Lipid A is the component responsible for the pathophysiology elicited by the parenteral use of LPS contaminated biologicals or reagents. These adverse effects are underscored by the over-activation of innate immune cells through pattern recognition receptors (PRRs), such as Toll-like receptors and specifically TLR4 homodimers in complex with myeloid differentiation factor (MD-2). LPS mediated over-induction of TLR4 signaling in monocytes promotes a generalized pro-inflammatory response leading to tissue damage, endotoxic shock, and death (VanAmersfoort et al. 2003, Chandler and Ernst, 2017).
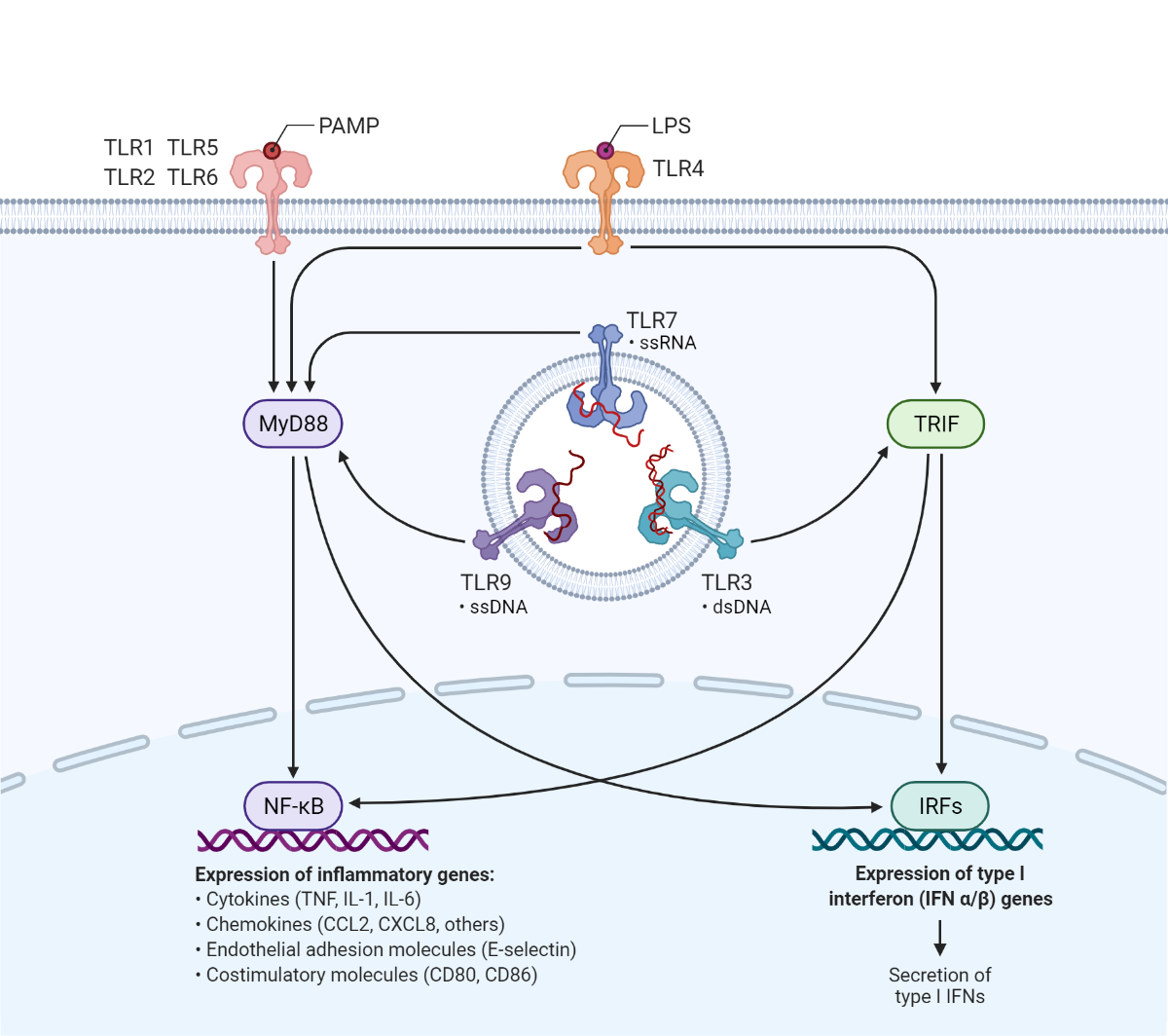
Reprinted from "TLR signaling pathway" by BioRender.com (2021). Retrieved from https://app.biorender.com/biorender-templates.
Pathogen-associated molecular patterns (PAMPs) are microbial-derived molecules recognized by the innate immune system. Toll-like receptors (TLRs) expressed by macrophages and dendritic cells interact with specific types of PAMPs, including LPS, leading to the production of inflammatory factors which promote anti-microbial immune responses.
Endotoxins in bioprocessing
Because of LPS-associated toxicity, biotherapeutics and any other biological intended for parenteral use in patients or animal models must meet established requirements to ensure their safety. For example, for humans and preclinical animal models, the endotoxin limit is 5 EU/kg (i.e., Endotoxin Units) for products administered parenterally over an hour. This limit is established by the United States Pharmacopoeia (USP) to ensure that parenteral administration of a product will not be pyrogenic or induce fever (Malyala and Singh, 2008). However, in practice, the goal is to maintain endotoxin levels as low as possible to prevent adverse effects. In the preclinical setting for both in vivo and in vitro testing, low endotoxin levels ensure that compounds are adequately evaluated for their actual biological effects and not those associated with endotoxin contamination.
To this end, during bioprocessing is not only essential to adopt effective strategies for LPS removal from biological products, but it is also equally important to maintain low LPS levels throughout the production workflow. This is not a trivial pursuit considering that LPS is not heat liable and resists conventional sterilizing methodologies (Magalhães et al. 2007). Additionally, LPS may be introduced during bioprocessing through contaminated water, buffers, cell culture media, glass or plasticware, and equipment.
Detecting and removing LPS
The FDA has approved two main methods for LPS detection: the rabbit pyrogen test and the Limulus Amoebocyte Lysate (LAL) assay. Nevertheless, LAL assays have become the standard for LPS detection in biologicals. Several LAL assays are commonly used, such as the gel-clot, turbidimetric, and chromogenic methods. In the most simple test, the LAL gel-clot assay, exposure of horseshoe crab blood cell lysate to endotoxin leads to coagulation, which is verified by visual inspection. The LAL gel-clot assay has a sensitivity of 0.03 EU/mL (Sandle 2016). The turbidimetric assay follows an endotoxin-induced increase in turbidity without gel-clot formation. In contrast, the chromogenic assay measures the hydrolysis of a chromogenic substrate (i.e., p-nitroaniline) in the presence of endotoxin contaminants. Overall, the turbidimetric and chromogenic LAL assays are faster and more accurate than the gel-clot method (Magalhães et al. 2007).
Endotoxin-protein interactions are the main challenge for LPS removal from biologics. Therefore, various methods have been developed to disrupt LPS-protein interactions or promote LPS binding to other substrates and away from the protein of interest. However, the specific method selected greatly depends on the properties of both the protein being purified and the contaminant LPS. Some of the techniques that may be leveraged for LPS removal from protein targets include ion-exchange chromatography, affinity adsorbents, gel filtration chromatography, sucrose gradient centrifugation, and Triton X-114 phase separation (Magalhães et al. 2007, Sandle 2016).
Reference
Chandler, C. E. & Ernst, R. K. Bacterial lipids: powerful modifiers of the innate immune response (2017) F1000 Res. doi: 10.12688/f1000research.11388.1.
de Oliveira Magalhães, P. et al. Methods of endotoxin removal from biological preparations: A review. Journal of Pharmacy and Pharmaceutical Sciences (2007). PMID: 17727802
Ding, X., Su, W. & Ding, X. Methods of Endotoxin Detection. Journal of Laboratory Automation (2015) doi:10.1177/2211068215572136.
Epstein, J., Kelly, C. E., Lee, M. M. & Donahoe, P. K. Effect of E. Coli endotoxin on mammalian cell growth and recombinant protein production. In Vitro Cellular & Developmental Biology (1990) doi:10.1007/BF02623686.
Malyala, P. & Singh, M. Endotoxin limits in formulations for preclinical research. Journal of Pharmaceutical Sciences (2008) doi:10.1002/jps.21152.
Ohto, U., Fukase, K., Miyake, K. & Shimizu, T. Structural basis of species-specific endotoxin sensing by innate immune receptor TLR4/MD-2. Proc. Natl. Acad. Sci. U. S. A. (2012) doi:10.1073/pnas.1201193109.
Sampath, V. P. Bacterial endotoxin-lipopolysaccharide; structure, function and its role in immunity in vertebrates and invertebrates. Agriculture and Natural Resources (2018) doi:10.1016/j.anres.2018.08.002.
Sandle, T. Endotoxin and pyrogen testing. in Pharmaceutical Microbiology (2016). doi:10.1016/b978-0-08-100022-9.00011-6.
Van Amersfoort, E. S., Van Berkel, T. J. C. & Kuiper, J. Receptors, mediators, and mechanisms involved in bacterial sepsis and septic shock. Clinical Microbiology Reviews (2003) doi:10.1128/CMR.16.3.379-414.2003.
- Like (2)
- Reply
-
Share



About Us · User Accounts and Benefits · Privacy Policy · Management Center · FAQs
© 2025 MolecularCloud